Nutritional Modulation of Epigenetic Changes Induced by Mycotoxins: A Biochemical Perspective for At-Risk Populations in Africa
-
David Chinonso Anih


Department of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba, Nigeria
Kayode Adebisi AroworaDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba, Nigeria
Kenneth Chinekwu UgwuokeDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba, Nigeria
Moses Adondua AbahDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba, Nigeria
Bilyaminu HabibuDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba, Nigeria
| Received 29 Jun, 2025 |
Accepted 12 Aug, 2025 |
Published 13 Aug, 2025 |
Mycotoxins, toxic secondary metabolites produced by fungi, are widespread contaminants of staple foods in Sub-Saharan Africa, contributing to acute toxicity and chronic diseases such as cancer, stunting, and immune dysfunction. Recent evidence has illuminated their capacity to induce epigenetic changes, including alterations in DNA methylation, histone modifications, and noncoding RNA expression, without modifying the DNA sequence. These disruptions, particularly when combined with nutritional deficiencies, can have long-lasting effects on gene regulation and disease susceptibility. This review synthesizes evidence from peer-reviewed studies published between 2007 and 2025, exploring how key nutrients modulate the epigenetic toxicity of common mycotoxins like aflatoxin B1, ochratoxin A, and fumonisin B1. A systematic literature search identified studies evaluating the biochemical and epigenetic responses to mycotoxin exposure and the mitigating roles of dietary factors such as folate, vitamin B12, choline, polyphenols, selenium, and short-chain fatty acids. These nutrients support critical pathways, including one-carbon metabolism, antioxidant defense, and gut microbiota modulation, which intersect with epigenetic regulation. Findings highlight that methyl donors can restore DNA methylation balance, antioxidants can prevent oxidative stress-induced histone and DNA damage, and fiber-rich diets promote microbial metabolites that act as epigenetic modulators. Additionally, exposure to multiple mycotoxins exerts synergistic effects that amplify epigenetic damage, especially in nutritionally vulnerable populations. This review also discusses emerging intersections between mycotoxins and biochemical domains such as neuroepigenetics, mitochondrial signaling, and microbiome-host interactions. Nutritional interventions ranging from diet diversification to bioactive-rich functional foods and probiotic supplementation emerge as promising, culturally adaptable strategies for mitigating these epigenetic threats. An integrative framework is proposed, linking food safety, public health policy, and personalized nutrition to strengthen resilience in affected communities. In conclusion, nutrition-sensitive interventions targeting epigenetic mechanisms offer a feasible and impactful approach to reducing the long-term health burden of mycotoxins in Africa. Future research should focus on biomarker discovery, field-based dietary trials, and AI-driven predictive models to inform context-specific mitigation strategies.
| Copyright © 2025 Anih et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Mycotoxins are toxic secondary metabolites produced by fungi that contaminate a wide range of agricultural commodities, including staple foods such as maize, groundnuts, and cassava, which are dietary mainstays for many populations in Africa1. The widespread presence of mycotoxins poses a significant threat to human and animal health, leading to various acute and chronic diseases2. Beyond their direct toxic effects, accumulating evidence suggests that mycotoxins can induce epigenetic modifications, altering gene expression without changing the underlying DNA sequence3. These epigenetic changes, including DNA methylation, histone modification, and non-coding RNA regulation, can have long-lasting impacts on cellular function and disease susceptibility, potentially contributing to developmental abnormalities, immune dysfunction, and increased cancer risk4,5.
Populations in Africa are particularly vulnerable to mycotoxin exposure due to several factors. These include suboptimal pre- and post-harvest storage practices that favor fungal growth and mycotoxin production, limited access to effective detoxification technologies, and often monotonous diets heavily reliant on susceptible crops6. Furthermore, co-exposure to multiple mycotoxins is common, potentially leading to synergistic or additive adverse health effects7. The intersection of high mycotoxin exposure and pre-existing nutritional deficiencies in many African communities creates a complex scenario where the impact of mycotoxin-induced epigenetic changes may be exacerbated8.
Nutrition plays a crucial role in maintaining cellular homeostasis and influencing epigenetic processes9. Dietary components, such as methyl donors (e.g., folate, vitamin B12, choline), antioxidants (e.g., vitamins A, C, E, selenium), and other bioactive compounds (e.g., polyphenols), can modulate the activity of enzymes involved in epigenetic regulation10. Therefore, understanding how specific nutritional factors can counteract or mitigate the epigenetic alterations induced by mycotoxins is of paramount importance for developing effective intervention strategies in at-risk African populations. This review aims to provide a biochemical perspective on the nutritional modulation of mycotoxin-induced epigenetic changes, highlighting potential dietary interventions that could offer protection against the long-term health consequences of mycotoxin exposure in Africa.
Figure 1 contrasts contaminated food with a nutrient-rich alternative. On the left, moldy corn kernels illustrate visible fungal spoilage and mycotoxin risk. On the right, a colorful plant-based meal features vegetables, legumes, grains, and fruits. The comparison emphasizes the importance of food safety and dietary quality in health protection.
SYSTEMATIC REVIEW APPROACH
This section outlines the systematic approach used to review the interplay between mycotoxins, epigenetics, and nutrition in African populations. Relevant literature was identified using electronic databases and specific keywords. Studies were selected based on their focus on epigenetic effects and nutritional interventions. Data extraction summarized key mycotoxins, epigenetic markers, study designs, and findings. A narrative synthesis grouped studies by mycotoxin type and dietary influence. Ethical considerations and research limitations were also addressed.
This review employed a systematic approach to identify and synthesize relevant literature on the interplay between mycotoxins, epigenetics, and nutrition, with a specific focus on the African context. Electronic databases, including PubMed, Scopus, Web of Science, and Google Scholar, were searched using a combination of keywords such as “Mycotoxins”, “Aflatoxin”, “Ochratoxin”, “Fumonisin”, “Zearalenone”, “Epigenetics”, “DNA methylation”, “Histone modification”, “Non-coding RNA”, “Nutrition”, “Diet”, “Vitamins”, “Minerals”, “Antioxidants”, and “Africa”. The search was limited to peer-reviewed articles and relevant reports published between 2007 and 2025 to ensure the inclusion of the most recent findings11.

|
This image contrasts food contamination and nutritional intervention. On the left side, corn kernels are visibly covered in mold, indicating spoilage and potential health risks due to mycotoxin contamination. On the right side, a vibrant, nutrient-rich meal is presented, consisting of fresh vegetables, grains, legumes, and fruits such as spinach, broccoli, chickpeas, brown rice, avocado, tomato slices, cucumber slices, and a bowl of mixed berries (strawberries, blueberries, and raspberries). This contrast highlights the importance of food safety and balanced nutrition in preventing health hazards
| Table 1: | Search strategy and databases used | |||
| Database | Keywords used | Years covered | Filter sapplied |
| PubMed | Mycotoxins, aflatoxin, ochratoxin, epigenetics, DNA methylation, nutrition, Africa |
2007-2025 | Peer-reviewed, full-text, English language11-17 |
| Scopus | Mycotoxins, DNA methylation, histone modification, polyphenols, antioxidants |
2007-2025 | Original research, African studies, nutritional focus11-17 |
| Web of Science | Fumonisin, zearalenone, non-coding RNA, vitamins, minerals, epigenetic markers |
2007-2025 | Relevance to African populations, nutritional interventions11-19 |
| Google Scholar | Mycotoxins, nutritional modulation, food safety, epigenetics, oxidative stress |
2007-2025 | Recent studies, high citation relevance, human/animal studies11-19 |
| The electronic databases (PubMed, Scopus, Web of Science, and Google Scholar) used to identify relevant studies. It includes search keywords related to mycotoxins, nutrition, and epigenetics, along with applied filters such as peer-reviewed articles and African population relevance | |||
Table 1 outlines the systematic approach utilized for literature search, specifying the databases (PubMed, Scopus, Web of Science, Google Scholar), relevant keywords, and filters applied to ensure the inclusion of high-quality, peer-reviewed studies focused on mycotoxins, nutrition, and epigenetics.
Studies were included if they investigated the epigenetic effects of mycotoxins in in vitro, in vivo, or human population studies, particularly those conducted in or relevant to African populations. Additionally, articles exploring the potential of nutritional interventions to modulate these epigenetic changes were considered. Exclusion criteria included studies focusing solely on the acute toxicological effects of mycotoxins without examining epigenetic mechanisms, studies unrelated to nutrition or epigenetics, and reviews or meta-analyses (although their reference lists were screened for potentially relevant primary research)12-19.
Table 2 provides an overview of studies examining the effects of mycotoxins on epigenetic mechanisms, including DNA methylation, histone modifications, and non-coding RNA alterations. It categorizes research based on mycotoxin type, study design, and the impact of nutritional interventions on mitigating epigenetic changes.
| Table 2: | Inclusion and exclusion criteria for literature review | |||
| Criteria | Description | Inclusion | Exclusion | Citations |
| Study type | Type of research evaluated for mycotoxin epigenetic impacts |
Original research articles (in vitro, in vivo, human studies) |
Review articles, editorials, or opinion pieces |
|
| Focus area | Central subject involving epigenetic mechanisms and nutritional modulation |
Studies addressing mycotoxins, epigenetics, and nutrition |
Studies focusing solely on toxicity or lacking nutritional/epigenetic relevance |
|
| Geographic relevance |
Applicability to African populations and agricultural contexts |
Research conducted in or relevant to African dietary and environmental conditions |
Studies from unrelated geographical or socio- economic settings |
Liu et al.20 |
| Publication period |
Ensures review includes current trends and up- to-date findings |
Studies published from 2007 to 2025 |
Articles published before 2020 or with an unverifiable date |
|
| Language | Comprehension and extraction of methodological detail |
English language studies |
Non-English papers without translation |
|
| Data quality | Assessed via transparency, statistical rigor, and proper control |
Peer-reviewed with robust methods and appropriate controls |
Poorly designed studies lacking reproducibility or statistical support |
|
| Intervention scope |
Specific nutritional interventions aimed at countering epigenetic effects |
Nutritional studies showing effects on DNA methylation, histone modifications, or ncRNA linked to mycotoxin exposure |
Studies lacking epigenetic endpoints or focused solely on acute toxicity |
|
| Accessibility | Availability of full-text data for extraction and evaluation |
Freely available or institutionally accessible full texts |
Abstract-only or inaccessible studies |
|
| Ethical compliance |
Consideration for ethical approval in human/animal studies |
Studies indicating IRB or ethical review approval where applicable |
Research without ethical documentation or approval details |
|
| That examine the epigenetic effects of mycotoxins, including DNA methylation, histone modifications, and non-coding RNA expression. It highlights experimental models, nutritional interventions, and research conducted in African populations, providing insight into the interplay between toxic exposure and dietary modulation | ||||
Data extraction involved summarizing the type of mycotoxin studied, the epigenetic markers investigated, the nutritional interventions explored, the study design, and the key findings. Special attention was paid to studies that directly examined the impact of nutrition on mycotoxin-induced epigenetic alterations. The quality of the included studies was assessed based on established criteria relevant to their design (e.g., appropriate controls, statistical power, clear reporting of methods and results)14-20. A narrative synthesis of the findings was conducted, grouping studies based on the specific mycotoxin and nutritional factor investigated15-20. Challenges and limitations in the existing literature, as well as potential areas for future research, were also identified16-20. The Vancouver citation style was used for referencing17-20. Ethical considerations related to research involving human participants and animal models were noted where applicable18-20. Efforts were made to identify studies that addressed the specific dietary patterns and nutritional deficiencies prevalent in at-risk populations in Africa19,20.
NUTRITIONAL MODULATION OF MYCOTOXIN-INDUCED EPIGENETIC CHANGES: MECHANISMS, EVIDENCE, AND IMPLICATIONS FOR AFRICA
This study explores how mycotoxins alter gene regulation through epigenetic modifications. It details how mycotoxins disrupt DNA methylation, histone modifications, and non-coding RNA expression, while highlighting the critical role of nutritional factors like methyl donors and antioxidants in counteracting these effects. The section also discusses the influence of the gut microbiota and the complexities of synergistic effects from multiple mycotoxins. Ultimately, it emphasizes the significant implications for African populations, advocating for nutritional interventions to mitigate mycotoxin-induced health risks.
| Table3: | Impact of methyl donors on mycotoxin-induced DNA methylation | |||
| Nutrient | Mechanism | Study model | Outcome | Citations |
| Folate | Provides methyl groups via the one-carbon metabolism pathway |
Animal and cell models | Modulates DNA methylation in response to mycotoxin exposure |
|
| Vitamin B12 | Cofactor in methionine synthase for methyl group recycling |
In vivo models | Protects against global DNA hypomethylation caused by mycotoxins |
Kok et al.28 Perlmutter et al.29 Milagro et al.30 |
| Choline | Donates methyl groups and supports SAM production |
In vitro hepatocyte studies |
Reduces mycotoxin-induced epigenetic gene silencing |
|
| Betaine | Supports remethylation of homocysteine to methionine |
Rodent models | Restores altered methylation patterns in liver cells |
|
| Polyphenols | Regulate DNA methyltransferase activity |
Various in vitro cancer and liver cell models |
Modulate methylation of tumor suppressor genes affected by AFB1 |
|
| This table summarizes experimental findings on key nutritional methyl donors, folate, Vitamin B12, choline, betaine, and polyphenols and their mechanisms in counteracting DNA methylation disruptions caused by mycotoxin exposure across various biological models | ||||
DNA methylation and nutritional modulators: The DNA methylation, the addition of a methyl group to cytosine residues in DNA, is a key epigenetic mark involved in gene silencing and genomic stability21. Mycotoxin exposure has been shown to disrupt DNA methylation patterns, leading to both hyper- and hypomethylation of specific genes22. Studies have indicated that Aflatoxin B1 (AFB1), a prevalent mycotoxin in Africa, can alter the methylation status of tumor suppressor genes and oncogenes, potentially contributing to hepatocarcinogenesis23,24.
Several nutritional factors play a critical role in maintaining proper DNA methylation.
Table 3 highlights the protective roles of dietary methyl donors against mycotoxin-induced epigenetic alterations. Folate and B12 maintain methyl group balance; choline and betaine support SAM production, while polyphenols modulate DNMT activity. These nutrients mitigate gene silencing and abnormal methylation patterns in liver and cancer models.
Methyl donors, such as folate, vitamin B12, choline, and betaine, are essential for the one-carbon metabolism pathway, which provides the methyl groups required for DNA methylation reactions25-30.
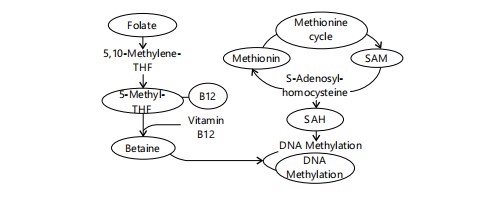
Figure 2 visually represents how dietary methyl donors regulate DNA methylation via the one-carbon metabolism pathway. Folate and betaine contribute methyl groups, while vitamin B12 acts as a cofactor in remethylation. These interactions sustain SAM synthesis, essential for DNA methyltransferase activity. Disruption by mycotoxins may impair this epigenetic balance.
Deficiencies in these nutrients, common in many African populations with limited dietary diversity, could exacerbate the effects of mycotoxins on DNA methylation26-30. Conversely, adequate intake of these methyl donors may offer some protection against mycotoxin-induced aberrant methylation patterns27-30. For instance, supplementation with folate has shown promise in modulating DNA methylation in various contexts28-30. Furthermore, dietary polyphenols found in fruits and vegetables, which are often lacking in sufficient quantities in the diets of at-risk populations in Africa, have also been shown to influence DNA methyltransferase (DNMT) activity, the enzymes responsible for DNA methylation29,30. Research exploring the direct impact of these nutritional interventions on mycotoxin-induced DNA methylation changes in African populations is crucial.
|
|
Histone modifications and dietary components: Histone modifications, including acetylation, methylation, phosphorylation, and ubiquitination of histone proteins around which DNA is wrapped, play a crucial role in regulating chromatin structure and gene accessibility31.
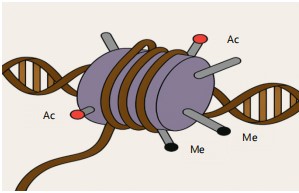
Figure 3 highlights the roles of histone acetylation and methylation in gene regulation. Acetylation typically relaxes chromatin to promote transcription, while methylation may activate or silence genes depending on context. These modifications are targets for dietary influence and are susceptible to disruption by mycotoxins such as ochratoxin A. Nutrients like SCFAs and phytochemicals can restore or modulate these marks to maintain epigenetic balance.
Mycotoxins have been implicated in altering histone modification patterns, thereby affecting gene expression32. For example, exposure to ochratoxin A (OTA), another significant mycotoxin contaminant, has been shown to influence histone acetylation and methylation in kidney cells33. These alterations can lead to dysregulation of genes involved in cell growth, apoptosis, and inflammation34.
Dietary components can influence histone modification enzymes, such as histone acetyltransferases (HATs) and Histone Deacetylases (HDACs)35. Short-Chain Fatty Acids (SCFAs), produced by the gut microbiota through the fermentation of dietary fiber, have been shown to inhibit HDAC activity36. Diets low in fiber, which is often the case in resource-limited settings in Africa, may compromise the production of SCFAs and thus the potential for HDAC inhibition37. Conversely, increased intake of fiber-rich foods could potentially counteract mycotoxin-induced alterations in histone acetylation.
| Table 4: | Dietary compounds influencing histone modification | |||
| Compound | Source | Effect on HAT/HDAC | Study outcome | Citations |
| Curcumin | Turmeric root | Inhibits HDAC activity | Restores histone acetylation balance and reduces mycotoxin- induced gene silencing |
|
| Butyrate | Fermentation of dietary fiber |
HDAC inhibitor | Enhances histone acetylation in colon and liver cells |
|
| Sulforaphane | Cruciferous vegetables (e.g., broccoli) |
Modulates HAT and HDAC enzymes |
Promotes gene expression through histone acetylation |
Fellows et al.37 |
| Ochratoxin A | Contaminated grains and coffee |
Alters histone methylation and acetylation |
Induces DNA damage and epigenetic dysregulation in renal cells |
Kaufman-Szymczyk et al.40 |
| Resveratrol | Red grapes, peanuts | Activates HAT, inhibits HDAC |
Regulates histone modifications and restores epigenetic balance |
|
| This table highlights key dietary compounds that modulate histone acetylation or methylation in the context of mycotoxin exposure, as evidenced by cellular and nutritional studies | ||||
Table 4 presents evidence from in vitro and nutritional studies showing how compounds like curcumin, butyrate, sulforaphane, and resveratrol modulate histone-modifying enzymes to influence gene expression38. Butyrate and sulforaphane act as HDAC inhibitors, enhancing histone acetylation, while curcumin and resveratrol exhibit dual roles affecting both HAT and HDAC activity. Conversely, ochratoxin A from contaminated grains disrupts histone acetylation and methylation patterns, contributing to epigenetic dysregulation.
Similarly, certain vitamins and minerals, as well as bioactive compounds like butyrate and sulforaphane found in cruciferous vegetables, can modulate histone modification39. Investigating the impact of these dietary factors on histone modification changes induced by prevalent mycotoxins in African populations is essential for identifying potential protective nutritional strategies40.
Non-coding RNAs and nutritional influence: Non-Coding RNAs (ncRNAs), such as microRNAs (miRNAs) and Long Non-Coding RNAs (lncRNAs), are important regulators of gene expression at the post-transcriptional level41. Emerging evidence suggests that mycotoxins can affect the expression profiles of various ncRNAs, contributing to their toxicological effects42. For instance, AFB1 exposure has been linked to altered expression of miRNAs involved in cell proliferation and apoptosis in liver cells43,44.
Nutritional factors have also been shown to modulate ncRNA expression45. Dietary fatty acids, vitamins, and polyphenols can influence the biogenesis and stability of miRNAs and lncRNAs46,47.
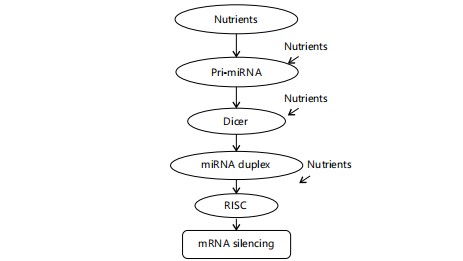
Figure 4 illustrates the nutrient-sensitive steps in miRNA biogenesis. Nutrients can modulate pri-miRNA transcription, Dicer-mediated processing, and miRNA duplex formation. These interactions collectively influence RISC assembly and gene silencing. Nutritional status therefore, critically impacts miRNA regulatory functions.
For example, omega-3 polyunsaturated fatty acids have been reported to alter miRNA expression in various tissues48. Given that the dietary intake of essential fatty acids and other micronutrients may be suboptimal in many African communities exposed to mycotoxins, understanding the interplay between nutrition, mycotoxins, and ncRNA regulation is critical49,50. Research exploring whether specific dietary interventions can restore or prevent mycotoxin-induced aberrant ncRNA expression in these populations is warranted.
|
Table 5 summarizes how nutritional components influence the expression of Non-Coding RNAs (ncRNAs) affected by exposure to major mycotoxins such as aflatoxin B1, ochratoxin A, and fumonisin B1. Specific nutrients, including polyphenols, omega-3 fatty acids, and vitamins A and D, have been shown to restore or modulate the expression of dysregulated miRNAs and lncRNAs. These interactions highlight the therapeutic potential of nutritional interventions in counteracting mycotoxin-induced epigenetic disturbances.
Antioxidants and epigenetic modulation: Oxidative stress, characterized by an imbalance between the production of Reactive Oxygen Species (ROS) and the capacity of the antioxidant defense system, is a common consequence of mycotoxin exposure51. The ROS can directly damage DNA and also indirectly influence epigenetic machinery52. Mycotoxins like AFB1 and OTA can induce oxidative stress, potentially leading to alterations in DNA methylation and histone modifications53,54.
Dietary antioxidants, including vitamins A, C, and E, selenium, and various phytochemicals, play a crucial role in neutralizing ROS and protecting against oxidative damage55. Adequate intake of these antioxidants may help mitigate the oxidative stress induced by mycotoxins and consequently reduce the associated epigenetic perturbations56.
Table 6 compiles key antioxidants such as vitamins C and E, selenium, polyphenols, and glutathione, and their capacity to reverse or prevent DNA methylation, histone modifications, and miRNA dysregulation. Experimental evidence from in vivo and in vitro studies underscores their potential in protecting against oxidative epigenetic toxicity caused by AFB1 and OTA exposure.
However, the dietary intake of these antioxidants is often low in at-risk populations in Africa57-60. Supplementation or dietary enrichment with antioxidant-rich foods could potentially offer protection against mycotoxin-induced epigenetic changes mediated by oxidative stress58-60. Studies investigating the impact of antioxidant interventions on epigenetic markers in individuals exposed to mycotoxins in Africa are needed.
| Table 5: | Mycotoxin and nutritional modulation of ncrna expression | |||
| Mycotoxin | Affected ncRNA | Nutrient modulator | Effect | Citations |
| Aflatoxin B1 | miR-34a | Polyphenols | Downregulation of miR-34a reversed; restored apoptosis balance |
|
| Aflatoxin B1 | miR-122, miR-192 |
Omega-3 fatty acids | Modulated miRNA expression, reduced inflammatory signals |
|
| Aflatoxin B1 | Multiple (miR-24, miR-125b) |
Vitamin A, D | Restored disrupted miRNA/lncRNA levels involved in hepatic detoxification |
|
| Aflatoxin B1 | miR-29a, lncRNA H19 |
Balanced diet (macro/micronutrients) |
Improved ncRNA homeostasis and reduced liver injury |
|
| Ochratoxin A | lncRNA MALAT1, miR-200c |
Polyphenols, vitamins | Counteracted oxidative stress and ncRNA misregulation |
El-Agamy50 |
| Fumonisin B1 | miR-21, miR-155 | Nutritionally enriched diets |
Attenuated pro-inflammatory ncRNA expression |
|
| The interplay between various mycotoxins and their impact on non-coding RNA (ncRNA) expression, with corresponding dietary modulators and their mitigating effects. Nutrients such as polyphenols, omega-3 fatty acids, and vitamins exhibit potential in restoring ncRNA balance disrupted by mycotoxins like aflatoxin B1, ochratoxin A, and fumonisin B1 | ||||
| Table 6: | Antioxidants that modulate epigenetic alterations | |||
| Antioxidant | Dietary source | Epigenetic effect | Evidence | Citations |
| Vitamin C and E | Citrus fruits, nuts, seeds |
Reduced AFB1-induced DNA methylation and histone modifications |
In vivo rodent model study showing epigenetic protecton |
Sagomba and Ndhlovu51 Lu et al.52 Alipoor et al.53 |
| Selenium and Vitamin E |
Nuts, cereals, eggs |
Combats mycotoxin- triggered oxidative and epigenetic damage |
Elemental synergy reduces histone and DNA methylation changes |
Jobe et al.54 Surai55 Albadrani et al.56 |
| Polyphenols | Fruits, vegetables, tea |
Prevented OTA-induced histone modification and DNA methylation |
Protective antioxidant pathway activation |
Tanvir et al.57 da Silva et al.58 Yang et al.59 |
| Glutathione | Spinach, avocado, asparagus |
Restored DNA methylation altered by AFB1 |
In vitro hepatic protection via methylation correction |
Loganathan and Doss60 |
| Flavonoids | Berries, dark chocolate |
Reversed mycotoxin-induced miRNA dysregulation and chromatin remodeling |
Epigenomic modulation through antioxidant action |
|
| General antioxidants |
Mixed plant-based diets |
Overall reduction of oxidative DNA damage and improved epigenetic stability |
Supported across reviews and experimental models |
|
| The role of various dietary antioxidants in mitigating epigenetic alterations caused by mycotoxin-induced oxidative stress. Evidence suggests that nutrients such as vitamins C and E, selenium, polyphenols, and glutathion e can reverse or prevent harmful modifications to DNA methylation and histone structure, thereby offering protective effects. The findings are supported by both in vivo and in vitro studies across diverse antioxidant sources | ||||
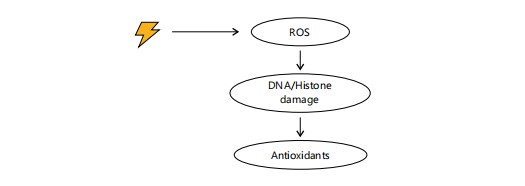
Figure 5 shows how mycotoxin-induced oxidative stress generates ROS that damage DNA and histones, contributing to epigenetic dysregulation59,60. These alterations can affect gene expression, potentially leading to adverse health outcomes. Antioxidants such as vitamins C and E, selenium, and polyphenols mitigate this oxidative and epigenetic damage by neutralizing ROS and restoring redox balance60. The figure supports the mechanisms described in this sub-section regarding antioxidant intervention in epigenetic modulation under oxidative stress60.
Gut microbiota and epigenetic interactions: The gut microbiota, a complex community of microorganisms residing in the gastrointestinal tract, plays a significant role in host health, including odulating immune responses and influencing metabolism61. Mycotoxins can disrupt the composition and function of the gut microbiota, leading to dysbiosis62. Conversely, the gut microbiota can metabolize some mycotoxins, potentially altering their bioavailability and toxicity63.
|
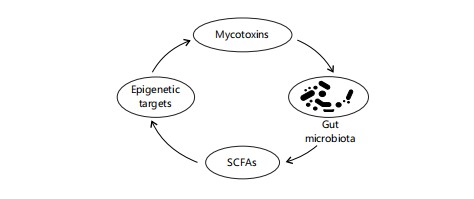
|
The bidirectional relationship between the gut microbiota and epigenetic modifications under the influence of dietary components and mycotoxin exposure. It shows how dysbiosis, SCFA production, and microbial metabolism of toxins modulate host epigenetic responses Emerging evidence suggests a bidirectional link between the gut microbiota and epigenetics64.
Figure 6 maps out the interplay between gut microbiota and host epigenetics in the context of mycotoxin exposure. It shows how mycotoxins disrupt microbial balance, while beneficial microbes produce metabolites like Short-Chain Fatty Acids (SCFAs) that modulate DNA methylation and histone acetylation. This interaction is diet-sensitive, highlighting the importance of fiber and prebiotics in regulating both microbial and epigenetic health.
Microbial metabolites, such as SCFAs, can influence histone modifications in the host65. Furthermore, the gut microbiota can indirectly affect DNA methylation through the production of metabolites involved in one-carbon metabolism66. Nutritional factors, particularly dietary fiber and prebiotics, can significantly impact the composition and function of the gut microbiota67. Diets rich in these components, which may be lacking in some African populations, can promote the growth of beneficial bacteria and the production of health-promoting metabolites68.
Table 7 outlines the role of microbial-derived metabolites such as butyrate, acetate, and propionate in regulating host epigenetic mechanisms. These metabolites influence histone acetylation and DNA methylation by acting as HDAC inhibitors or providing substrates for methylation. Their production is modulated by dietary intake of fibers and prebiotics, supporting gut-epigenome homeostasis.
| Table 7: | Microbial metabolites involved in epigenetic regulation | |||
| Metabolite | Microbial source | Epigenetic target | Dietary driver | Citations |
| Butyrate prausnitzii and |
Faecalibacterium (HDACs) Roseburia spp. |
Histone deacetylases (prebiotics) |
Dietary fiber Rudrapal et al.62 |
|
| Acetate muciniphila |
Bacteroides spp. |
DNA methyltransferases (DNMTs), HDACs |
Resistant starch, oligosaccharides |
|
| Propionate | Bacteroides spp. |
HDAC inhibition, miRNA regulation |
Whole grains, legumes |
|
| Indole derivatives | Clostridium spp. |
Aryl hydrocarbon receptor (AhR) modulation |
Tryptophan-rich diets | Holmes et al.69 |
| Secondary bile acids | Bacteroides spp. |
Histone acetylation, nuclear receptor signaling |
High-fat diet, bile salt metabolism |
|
| Lactate modification |
Lactobacillus spp. |
HDAC inhibition, histone | Fermented foods, | |
| Phenylacetic acid | Clostridium spp. and |
Histone methylation | Protein-rich diets | |
| Aflatoxin- degrading enzymes |
Lactobacillus plantarum | Modulates toxin-induced epigenetic stress |
Probiotic supplementation |
|
| This table summarizes key microbial metabolites produced in the gut that influence host epigenetic mechanisms. It highlights the microbial taxa responsible for generating each metabolite, their respective epigenetic targets (including HDACs, DNMTs, histone modifications, and miRNAs), and the dietary components that drive their production. The interplay of these factors underpins a dynamic microbe-diet-epigenome axis relevant to health and disease | ||||
Investigating how dietary interventions aimed at modulating the gut microbiota can influence mycotoxin-induced epigenetic changes in at-risk African populations represents a promising area of research68,69.
Synergistic effects of multiple mycotoxins and nutrition: In real-world scenarios, particularly in Africa, exposure to multiple mycotoxins is common70. These co-exposures can lead to synergistic or additive toxic effects, which may also extend to epigenetic modifications71. For example, combined exposure to AFB1 and fumonisins has been shown to have more severe health consequences than exposure to either mycotoxin alone72.
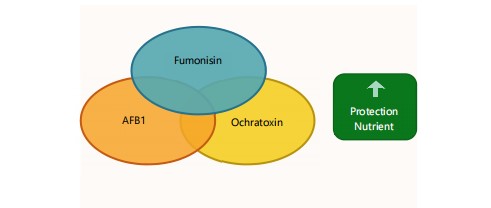
Figure 7 visualizes the enhanced toxicity of mycotoxin mixtures (e.g., AFB1+fumonisin) and their epigenetic impacts73. Nutritional interventions (polyphenols, probiotics, micronutrients) disrupt synergistic damage by mitigating oxidative stress and genotoxicity. The model highlights how nutrient deficiencies exacerbate vulnerability in co-exposure scenarios, while dietary diversification offers protection. It contextualizes Table 8 data within real-world exposure contexts in Africa.
The nutritional status of individuals can also influence the combined effects of multiple mycotoxins on epigenetic regulation74. Deficiencies in essential nutrients may render individuals more susceptible to the adverse effects of co-exposure. Conversely, adequate or enhanced intake of specific nutrients may offer broader protection against the epigenetic disruption caused by multiple mycotoxins75.
Table 8 catalyses evidence on mycotoxin co-exposure synergism (e.g., aflatoxin+fumonisin nephrotoxicity) and nutritional countermeasures75,76. Deficiencies in selenium/zinc exacerbate epigenetic dysregulation, while antioxidant-rich diets reduce DNA damage76,77. African field studies confirm growth retardation from mixed exposures in maize-dependent populations77. Data advocate integrative nutrition (e.g., probiotic/isolavone supplementation) to break toxicity cycles78.
|
| Table 8: | Combined effects of mycotoxins and nutritional status | |||
| Mycotoxin combo | Effect | Nutritional modulator | Study design | Citations |
| Aflatoxin+Fumonisin | Synergistic intestinaland liver toxicity | Antioxidant-rich diets | In vitro and animal models |
|
| Multiple mycotoxins(AFB1, DON, ZEN) | Enhanced epithelial damage and immune suppression |
Micronutrients (Se, Zn, Vit A) |
In vitro and human exposure data |
|
| Aflatoxin+Zearalenone | Endocrine disruption and genotoxicity |
Isoflavones and probiotics |
Animal experiments |
|
| Co-occurring fumonisin, aflatoxin, ochratoxin |
Growth retardation, gut permeability alteration |
Dietary diversity and protein adequacy |
Field studies in children |
|
| Mixed mycotoxin exposure in maize |
Cumulative dietary toxicity |
Food processing (fermentation, nixtamalization) |
Dietary surveys and food analysis |
|
| Aflatoxin+Ochratoxin A | Nephrotoxicity and oxidative stress |
Polyphenols and flavonoids |
Animal models and mechanistic studies |
|
| Multiple mycotoxins in infants |
Compromised immunity and micronutrient |
Infant complementary feeding strategies |
Cross-sectional and cohort studies |
|
| General co-exposure | Synergistic toxicity and malnutrition interaction |
Nutritional counselling and interventions |
Review and meta-analysis | |
| The interactive effects of combined mycotoxin exposures and nutritional status. It details specific mycotoxin combinations, their toxicological outcomes, modulating nutritional factors, and the nature of supporting studies. These findings underscore the importance of integrative nutritional strategies to mitigate mycotoxin-related health risks, especially in vulnerable populations across Sub-Saharan Africa | ||||
Research specifically examining the nutritional modulation of epigenetic changes induced by mixtures of mycotoxins, relevant to the African context, is crucial for developing effective risk mitigation strategies79. This research should consider the common co-occurrence of mycotoxins in staple foods and the prevalent nutritional deficiencies in these populations.
IMPLICATIONS FOR AT-RISK POPULATIONS IN AFRICA
The findings discussed in this review have significant implications for at-risk populations in Africa who face chronic exposure to mycotoxins and often experience nutritional deficiencies80. Understanding how nutrition can modulate mycotoxin-induced epigenetic changes opens avenues for developing targeted dietary interventions to mitigate the long-term health risks associated with mycotoxin exposure81.

|
| Table 9: | Recommendations for nutritional interventions | |||
| Strategy | Nutrient focus | Implementation approach | Target population | Citations |
| Functional food | Anti-epigenetic | Encourage consumption | Populations at risk | Turner et al.84 |
| promotion | bioactives | of biofortified and functional foods |
of aflatoxin exposure | Nji et al.85 Visser et al.86 |
| Nutrition-sensitive food safety education |
Mycotoxin awareness, micronutrients |
Integrate food safety into nutrition education programs |
Women, caregivers, and school-age children |
Kimanya et al.87 Ingenbleek et al.88 Wild and Gong89 |
| Community-based hygiene interventions |
Sanitation-linked nutrient retention |
Local programs addressing water, sanitation, and hygiene (WASH) |
Underserved rural communities |
|
| Policy-driven public health campaigns |
Nutrient-enabling environments |
National aflatoxin mitigation strategies and food system reforms |
Policymakers, educators, and food vendors |
|
| Risk communication and systems approach |
Environmental nutrition interaction |
Holistic assessment of environmental and dietary toxins |
General African populations in hotspot regions |
|
| This table presents strategic nutritional interventions to mitigate mycotoxin exposure risks across African communities. Dietary diversification, functional food intake, food safety education and hygiene-centered programs are recommended for vulnerable populations. Policy campaigns and systems-based communication target broader public health infrastructure and environmental factors | ||||
Figure 8 proposes an integrated strategy to address mycotoxin exposure in African populations. Food safety practices reduce contamination in staples81, while nutrition education promotes protective diets rich in methyl donors and antioxidants82. Community-driven hygiene interventions enhance nutrient retention82,83, and research identifies epigenetic biomarkers for intervention efficacy83. The model aligns with Table 9 evidence-based recommendations for vulnerable groups.
Promoting dietary diversity, increasing the intake of fruits, vegetables, and whole grains rich in methyl donors, antioxidants, and fiber, and addressing micronutrient deficiencies are crucial steps83.
Table 9 synthesizes intervention strategies to disrupt mycotoxin-induced epigenetic damage in Africa. Dietary diversification combats micronutrient deficiencies linked to susceptibility83,84, while functional foods (e.g., biofortified crops) deliver anti-epigenetic bioactives84. Policy-driven food safety campaigns and WASH programs address environmental determinants85, and systems-based risk communication enables holistic responses86.
|
Furthermore, research is needed to identify specific nutritional biomarkers that can indicate susceptibility to mycotoxin-induced epigenetic damage and to monitor the effectiveness of dietary interventions87.
Culturally appropriate and sustainable food-based strategies should be prioritized88. Public health initiatives aimed at improving food safety practices, reducing mycotoxin contamination in staple crops, and promoting nutrition education are essential components of a comprehensive approach to protect the health of these vulnerable populations89. Future research should focus on conducting well-designed intervention studies in African settings to evaluate the efficacy of specific nutritional strategies in modulating mycotoxin-related epigenetic alterations and improving health outcomes.
INTEGRATIVE PERSPECTIVES ON EMERGING BIOCHEMICAL INTERACTIONS AND NUTRITIONAL EPIGENETICS
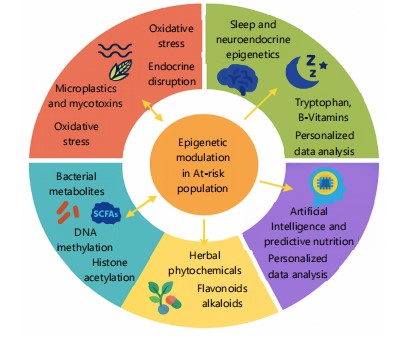
Recent advances in nutritional epigenetics have expanded our understanding of how biochemical signals interact with environmental toxins to modulate gene expression. Mycotoxins are increasingly being considered alongside other emerging pollutants such as microplastics, which may similarly disrupt epigenetic homeostasis through oxidative stress, mitochondrial dysfunction, and interference with hormone signaling pathways90. These pollutants, including mycotoxins, can influence systemic biochemical networks, with cumulative effects on metabolic, immune, and neuroendocrine systems.
A growing body of evidence indicates that neurobiological rhythms, particularly those governing sleep and circadian cycles, are under both nutritional and e pigenetic regulation91. Mycotoxins have been associated with altered sleep architecture and endocrine function, potentially via modulation of melatonin pathways and DNA methylation enzymes. Dietary modulation through polyphenols, methyl donors, and specific amino acids may provide a corrective mechanism to restore disrupted neuroepigenetic circuits.
Moreover, bacterial-derived metabolites are emerging as potent biochemical mediators that influence host immunity, neurotransmitter regulation, and gene expression through epigenetic remodeling92. These findings emphasize the potential of microbiome-targeted nutritional strategies such as the use of prebiotics, probiotics, and postbiotics to counteract mycotoxin-induced epigenetic alterations.
Parallel insights from herbal biochemistry offer another promising intervention pathway. Certain phytochemicals from traditional herbal medicines possess epigenetic activity, including inhibition of Histone Deacetylases (HDACs) and DNA Methyltransferases (DNMTs), thereby modulating gene expression in a manner that can mitigate mycotoxin toxicity93. The integration of nutraceuticals with known molecular targets could strengthen community-based dietary interventions.
Finally, Artificial Intelligence (AI) is beginning to play a pivotal role in biochemical research, offering computational models to predict nutrient toxin epigenome interactions94. The AI-based platforms can help identify population-specific dietary strategies that optimally counteract the epigenetic burden of mycotoxins, accounting for genetic polymorphisms, metabolic profiles, and environmental exposures. Future research in African populations should incorporate these novel perspectives to enhance the effectiveness of nutrition-sensitive interventions.
Figure 9 illustrates how diverse biochemical and environmental factors influence epigenetic modulation in at-risk populations. Key contributors include microplastics, gut microbiota, sleep regulation, herbal phytochemicals, and AI-guided nutrition. Arrows represent directional influences on DNA methylation, histone acetylation, and overall epigenetic stability.
CONCLUSION
Mycotoxin exposure poses a significant epigenetic and public health challenge in Sub-Saharan Africa, especially among nutritionally vulnerable populations. This review highlights the pivotal role of nutrition in modulating mycotoxin-induced alterations in DNA methylation, histone modifications, and noncoding RNA expression. Key dietary components, including methyl donors, antioxidants, and fiber, offer promising protective mechanisms against these epigenetic disruptions. Integrating nutrition-sensitive strategies with food safety interventions can mitigate long-term health risks. Culturally appropriate dietary interventions and public health education are essential for sustainable impact. Future research should prioritize biomarker discovery and targeted trials to inform evidence-based policies.
SIGNIFICANCE STATEMENT
This manuscript highlights a critical yet often overlooked dimension of public health in Sub-Saharan Africa, the intersection between chronic mycotoxin exposure and nutrition-driven epigenetic regulation. By synthesizing current evidence on how key dietary components such as methyl donors, antioxidants, and microbiota modulators influence gene expression and mitigate toxin-induced epigenetic disruptions, the study presents a compelling framework for preventive health. It emphasizes the potential of nutrition as a cost-effective, culturally appropriate strategy to reduce the long-term health burden associated with mycotoxins. The findings offer valuable insights for integrating nutritional interventions into food safety policies and population health programs.
REFERENCES
- Imade, F., E.M. Ankwasa, H. Geng, Sana Ullah and T. Ahmad et al., 2021. Updates on food and feed mycotoxin contamination and safety in Africa with special reference to Nigeria. Mycology, 12: 245-260.
- Ezekiel, C.N., W.A. Abia, D. Braun, B. Šarkanj and K.I. Ayeni et al., 2022. Mycotoxin exposure biomonitoring in breastfed and non-exclusively breastfed Nigerian children. Environ. Int., 158.
- Mungamuri, S.K. and V.A. Mavuduru, 2020. Role of epigenetic alterations in aflatoxin-induced hepatocellular carcinoma. Liver Cancer Int., 1: 41-50.
- Molina-Hernandez, J.B., C.D. Grande-Tovar, L. Neri, J. Delgado-Ospina, M. Rinaldi, G.A. Cordero-Bueso and C. Chaves-López, 2025. Enhancing postharvest food safety: The essential role of non-thermal technologies in combating fungal contamination and mycotoxins. Front. Microbiol., 16.
- Wenndt, A., F. Mutua, D. Grace, L.F. Thomas and E. Lambertini, 2023. Quantitative assessment of aflatoxin exposure and hepatocellular carcinoma (HCC) risk associated with consumption of select Nigerian staple foods. Front. Sustainable Food Syst., 7.
- Obonyo, M.A. and E.N. Salano, 2018. Perennial and seasonal contamination of maize by aflatoxins in Eastern Kenya. Int. J. Food Contam., 5.
- Mahato, D.K., K.E. Lee, M. Kamle, S. Devi, K.N. Dewangan, P. Kumar and S.G. Kang, 2019. Aflatoxins in food and feed: An overview on prevalence, detection and control strategies. Front. Microbiol., 10.
- Aasa, A.O., F.F. Fru, O.A. Adelusi, S.A. Oyeyinka and P.B. Njobeh, 2023. A review of toxigenic fungi and mycotoxins in feeds and food commodities in West Africa. World Mycotoxin J., 16: 33-48.
- Pazmiño, A.Y.G. and B.E.C. García, 2025. Nutritional epigenetics. EPRA Int. J. Multidiscip. Res., 11: 137-141.
- Cory, H., S. Passarelli, J. Szeto, M. Tamez and J. Mattei, 2018. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr., 5.
- Eskola, M., G. Kos, C.T. Elliott, J. Hajšlová, S. Mayar and R. Krska, 2020. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr., 60: 2773-2789.
- Nsabiyumva, G., C.K. Mutegi, J.M. Wagacha, A.B. Mohamed and N.K. Njeru et al., 2023. Aflatoxin contamination of maize and groundnut in Burundi: Distribution of contamination, identification of causal agents and potential biocontrol genotypes of Aspergillus flavus. Front. Microbiol., 14.
- Watson, S., S.E. Moore, M.K. Darboe, G. Chen and Y.K. Tu et al., 2018. Impaired growth in rural Gambian infants exposed to aflatoxin: A prospective cohort study. BMC Public Health, 18.
- Agriopoulou, S., E. Stamatelopoulou and T. Varzakas, 2020. Advances in analysis and detection of major mycotoxins in foods. Foods, 9.
- Ntsama, I.S.B., C. Frazzoli, G.B. Pouokam and V. Colizzi, 2023. Occurrence and dietary risk assessment of mycotoxins in most consumed foods in Cameroon: Exploring current data to understand futures challenges. Foods, 12.
- Hattle, M., D.L. Burke, T. Trikalinos, C.H. Schmid, Y. Chen, D. Jackson and R.D. Riley, 2022. Multivariate meta-analysis of multiple outcomes: Characteristics and predictors of borrowing of strength from Cochrane reviews. Syst. Rev., 11.
- Ezekiel, C.N., K.I. Ayeni, M.O. Akinyemi, M. Sulyok and O.A. Oyedele et al., 2021. Dietary risk assessment and consumer awareness of mycotoxins among household consumers of cereals, nuts and legumes in North-Central Nigeria. Toxins, 13.
- Thompson, P.B., 2025. Perspective: Ethical considerations of animal-sourced foods. Anim. Front., 15: 91-93.
- Ahmad, A. and J.H. Yu, 2017. Occurrence, toxicity and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health., 14.
- Liu, Y., J.H.G. Yamdeu, Y.Y. Gong and C. Orfila, 2020. A review of postharvest approaches to reduce fungal and mycotoxin contamination of foods. Compr. Rev. Food Sci. Food Saf., 19: 1521-1560.
- Kapri, A., D. Singh and S.K. Onteru, 2025. Deciphering aflatoxin B1 affected critical molecular pathways governing cancer: A bioinformatics study using CTD and PANTHER databases. Mycotoxin Res., 41: 93-111.
- Chu, Y.J., H.I. Yang, H.C. Wu, M.H. Lee and J. Liu et al., 2018. Aflatoxin B1 exposure increases the risk of hepatocellular carcinoma associated with hepatitis C virus infection or alcohol consumption. Eur. J. Cancer, 94: 37-46.
- Wu, H.C., Q. Wang, H.I. Yang, W.Y. Tsai, C.J. Chen and R.M. Santella, 2013. Global DNA methylation in a population with aflatoxin B1 exposure. Epigenetics, 8: 962-969.
- Fasullo, M., 2018. Cellular Responses to Aflatoxin-Associated DNA Adducts. In: DNA Repair-An Update, Mognato, M. (Ed.), IntechOpen, London, United Kingdom, ISBN: 978-1-83880-784-9.
- Serefidou, M., A.V. Venkatasubramani and A. Imhof, 2019. The impact of one carbon metabolism on histone methylation. Front. Genet., 10.
- Gachara, G., R. Suleiman, B. Kilima, M. Taoussi and S. El Kadili et al., 2024. Pre- and post-harvest aflatoxin contamination and management strategies of Aspergillus spoilage in East African Community maize: Review of etiology and climatic susceptibility. Mycotoxin Res., 40: 495-517.
- Amenyah, S.D., C.F. Hughes, M. Ward, S. Rosborough and J. Deane et al., 2020. Influence of nutrients involved in one-carbon metabolism on DNA methylation in adults-A systematic review and meta-analysis. Nutr. Rev., 78: 647-666.
- Kok, D.E., R.C. Richmond, M. Adriaens, C.T. Evelo and D. Ford et al., 2022. Impact of in utero folate exposure on DNA methylation and its potential relevance for later-life health-evidence from mouse models translated to human cohorts. Mol. Nutr. Food Res., 66.
- Perlmutter, A., J.S. Bland, A. Chandra, S.S. Malani, R. Smith, T.L. Mendez and V.B. Dwaraka, 2024. The impact of a polyphenol-rich supplement on epigenetic and cellular markers of immune age: A pilot clinical study. Front. Nutr., 11.
- Milagro, F.I., M.L. Mansego, C. de Miguel and J.A. Martínez, 2013. Dietary factors, epigenetic modifications and obesity outcomes: Progresses and perspectives. Mol. Aspects Med., 34: 782-812.
- Meir, A.Y., M. Keller, A. Hoffmann, E. Rinott and G. Tsaban et al., 2023. The effect of polyphenols on DNA methylation-assessed biological age attenuation: The DIRECT PLUS randomized controlled trial. BMC Med., 21.
- Tiffon, C., 2018. The impact of nutrition and environmental epigenetics on human health and disease. Int. J. Mol. Sci., 19.
- Aricthota, S., P.P. Rana and D. Haldar, 2022. Histone acetylation dynamics in repair of DNA double-strand breaks. Front. Genet., 13.
- Ozawa, S., R. Ojiro, Q. Tang, X. Zou, M. Jin, T. Yoshida and M. Shibutani, 2024. Involvement of multiple epigenetic mechanisms by altered DNA methylation from the early stage of renal carcinogenesis before proliferative lesion formation upon repeated administration of ochratoxin A. Toxicology, 506.
- Sasaki, K. and T. Masaki, 2025. Epigenetic histone modifications in kidney disease and epigenetic memory. Clin. Exp. Nephrol.
- Franzago, M., D. Santurbano, E. Vitacolonna and L. Stuppia, 2020. Genes and diet in the prevention of chronic diseases in future generations. Int. J. Mol. Sci., 21.
- Fellows, R., J. Denizot, C. Stellato, A. Cuomo and P. Jain et al., 2018. Microbiota derived short chain fatty acids promote histone crotonylation in the colon through histone deacetylases. Nat. Commun., 9.
- Martin-Gallausiaux, C., L. Marinelli, H.M. Blottière, P. Larraufie and N. Lapaque, 2021. SCFA: Mechanisms and functional importance in the gut. Proc. Nutr. Soc., 80: 37-49.
- Yuille, S., N. Reichardt, S. Panda, H. Dunbar and I.E. Mulder, 2018. Human gut bacteria as potent class I histone deacetylase inhibitors in vitro through production of butyric acid and valeric acid. PLoS ONE, 13.
- Kaufman-Szymczyk, A., G. Majewski, K. Lubecka-Pietruszewska and K. Fabianowska-Majewska, 2015. The role of sulforaphane in epigenetic mechanisms, including interdependence between histone modification and DNA methylation. Int. J. Mol. Sci., 16: 29732-29743.
- Dashwood, R.H. and E. Ho, 2008. Dietary agents as histone deacetylase inhibitors: Sulforaphane and structurally related isothiocyanates. Nutr. Rev., 66: S36-S38.
- Abdelmonem, B.H., L.T. Kamal, L.W. Wardy, M. Ragheb, M.M. Hanna, M. Elsharkawy and A. Abdelnaser, 2025. Non-coding RNAs: Emerging biomarkers and therapeutic targets in cancer and inflammatory diseases. Front. Oncol., 15.
- Marrone, A.K., V. Tryndyak, F.A. Beland and I.P. Pogribny, 2016. MicroRNA responses to the genotoxic carcinogens aflatoxin B1 and benzo[a] pyrene in human HepaRG cells. Toxicol. Sci., 149: 496-502.
- Lizarraga, D., S. Gaj, K.J. Brauers, L. Timmermans, J.C. Kleinjans and J.H.M. van Delft, 2012. Benzo[a]pyrene-induced changes in microRNA-mRNA networks. Chem. Res. Toxicol., 25: 838-849.
- Hua, Z., R. Liu, Y. Chen, G. Liu and C. Li et al., 2021. Contamination of aflatoxins induces severe hepatotoxicity through multiple mechanisms. Front. Pharmacol., 11.
- Lee, W., 2024. MicroRNAs and other non-coding RNAs as regulators, biomarkers, and therapeutic targets. Int. J. Mol. Sci., 25.
- Wyatt, M.M., L.W. Huff, M.H. Nelson, L.R. Neal and A.R. Medvec et al., 2023. Augmenting TCR signal strength and ICOS costimulation results in metabolically fit and therapeutically potent human CAR Th17 cells. Mol. Ther., 31: 2120-2131.
- Gareev, I., O. Beylerli, T. Ilyasova and C. Wang, 2024. Polyphenol-mediated modulation of non-coding RNAs: A new therapeutic approach for hypertension-A review. Curr. Hypertens. Rev., 20: 127-140.
- Ishihara, T., M. Yoshida and M. Arita, 2019. Omega-3 fatty acid-derived mediators that control inflammation and tissue homeostasis. Int. Immunol., 31: 559-567.
- El-Agamy, D.S., 2010. Comparative effects of curcumin and resveratrol on aflatoxin B1-induced liver injury in rats. Arch. Toxicol., 84: 389-396.
- Sagomba, E. and E. Ndhlovu, 2025. Food System Transformation for Safe and Nutritious Diets in Africa. In: African Food Systems: Rethinking Prospects for Continental Sustainable Transformation, Ndhlovu, E. (Ed.), Springer Nature, Switzerland, ISBN: 978-3-031-90823-1, pp: 241-265.
- Lu, Q., A. Wu, L. Tesmer, D. Ray, N. Yousif and B. Richardson, 2007. Demethylation of CD40LG on the inactive X in T cells from women with lupus. J. Immunol., 179: 6352-6358.
- Alipoor, B., S. Nikouei, F. Rezaeinejad, S.N. Malakooti-Dehkordi, Z. Sabati and H. Ghasemi, 2021. Long non-coding RNAs in metabolic disorders: Pathogenetic relevance and potential biomarkers and therapeutic targets. J. Endocrinol. Invest., 44: 2015-2041.
- Jobe, M.C., D.M.N. Mthiyane, P.V. Dludla, S.E. Mazibuko-Mbeje, D.C. Onwudiwe and M. Mwanza, 2023. Pathological role of oxidative stress in aflatoxin-induced toxicity in different experimental models and protective effect of phytochemicals: A review. Molecules, 28.
- Surai, P.F., 2015. Silymarin as a natural antioxidant: An overview of the current evidence and perspectives. Antioxidants, 4: 204-247.
- Albadrani, G.M., A.E. Altyar, O.A. Kensara, M.A.M. Haridy and M.S. Zaazouee et al., 2024. Antioxidant, anti-inflammatory, and anti-DNA damage effects of carnosic acid against aflatoxin B1-induced hepatic, renal, and cardiac toxicities in rats. Toxicol. Res., 13.
- Tanvir, I., A. Hassan and F. Albeladi, 2022. DNA methylation and epigenetic events underlying renal cell carcinomas. Cureus, 14.
- da Silva, E.O., A.P.F.L. Bracarense and I.P. Oswald, 2018. Mycotoxins and oxidative stress: Where are we? World Mycotoxin J., 11: 113-134.
- Yang, J.Y., J.J. Du, Y.F. Zhang and Y.X. Li, 2022. Vitamin E and selenium partially prevent cytotoxicity, oxidative stress and DNA damage induced by T-2 toxin in bovine Leydig cells. Theriogenology, 189: 255-261.
- Loganathan, T., and C.G.P. Doss, 2023. Non-coding RNAs in human health and disease: Potential function as biomarkers and therapeutic targets. Funct. Integr. Genomics, 23.
- Jalouli, M., M. Ataur Rahman, P. Biswas, Hasanur Rahman and A.H. Harrath et al., 2025. Targeting natural antioxidant polyphenols to protect neuroinflammation and neurodegenerative diseases: A comprehensive review. Front. Pharmacol., 16.
- Rudrapal, M., S.J. Khairnar, J. Khan, Abdulaziz Bin Dukhyil and M.A. Ansari et al., 2022. Dietary polyphenols and their role in oxidative stress-induced human diseases: Insights into protective effects, antioxidant potentials and mechanism(s) of action. Front. Pharmacol., 13.
- Owumi, S.E., C.E. Irozuru, U.O. Arunsi and A.K. Oyelere, 2022. Caffeic acid protects against DNA damage, oxidative and inflammatory mediated toxicities, and upregulated caspases activation in the hepatorenal system of rats treated with aflatoxin B1. Toxicon, 207: 1-12.
- Afzaal, M., F. Saeed, Y.A. Shah, M. Hussain and R. Rabail et al., 2022. Human gut microbiota in health and disease: Unveiling the relationship. Front. Microbiol., 13.
- Choi, H., Y. Garavito-Duarte, A.R. Gormley and S.W. Kim, 2025. Aflatoxin B1: Challenges and strategies for the intestinal microbiota and intestinal health of monogastric animals. Toxins, 17.
- Guerre, P., 2020. Mycotoxin and gut microbiota interactions. Toxins, 12.
- Zhang, Q., Y. Liu, Y. Li, G. Bai and J. Pang et al., 2025. Implications of gut microbiota-mediated epigenetic modifications in intestinal diseases. Gut Microbes, 17.
- Tian, S. and M. Chen, 2024. Global research progress of gut microbiota and epigenetics: Bibliometrics and visualized analysis. Front. Immunol., 15.
- Holmes, Z.C., M.M. Villa, H.K. Durand, S. Jiang and E.P. Dallow et al., 2022. Microbiota responses to different prebiotics are conserved within individuals and associated with habitual fiber intake. Microbiome, 10.
- Oyedele, O.A., C.N. Ezekiel, M. Sulyok, M.C. Adetunji, B. Warth, O.O. Atanda and R. Krska, 2017. Mycotoxin risk assessment for consumers of groundnut in domestic markets in Nigeria. Int. J. Food Microbiol., 251: 24-32.
- Hoffmann, V., K. Jones and J.L. Leroy, 2018. The impact of reducing dietary aflatoxin exposure on child linear growth: A cluster randomised controlled trial in Kenya. BMJ Global Health, 3.
- Paulo, L.S., G.M. Bwire, K. Klipstein-Grobusch, A. Kamuhabwa and G. Kwesigabo et al., 2023. Urbanization gradient, diet, and gut microbiota in Sub-Saharan Africa: A systematic review. Front. Microbiomes, 2.
- Li, D., Y. Li, S. Yang, J. Lu, X. Jin and M. Wu, 2022. Diet-gut microbiota-epigenetics in metabolic diseases: From mechanisms to therapeutics. Biomed. Pharmacother., 153.
- Pastor, L., A. Vettorazzi, J. Campión, P. Cordero and A.L. de Cerain, 2016. Gene expression kinetics of renal transporters induced by ochratoxin A in male and female F344 rats. Food Chem. Toxicol., 98: 169-178.
- Divella, R., A. Daniele, E. Savino and A. Paradiso, 2020. Anticancer effects of nutraceuticals in the mediterranean diet: An epigenetic diet model. Cancer Genomics Proteomics, 17: 335-350.
- Sagy, N., N. Meyrom, P. Beckerman, O. Pleniceanu and D.Z. Bar, 2024. Kidney-specific methylation patterns correlate with kidney function and are lost upon kidney disease progression. Clin. Epigenet., 16.
- Kamala, A., M. Kimanya, C. Lachat, L. Jacxsens and G. Haesaert et al., 2017. Risk of exposure to multiple mycotoxins from maize-based complementary foods in Tanzania. J. Agric. Food Chem., 65: 7106-7114.
- Gruber-Dorninger, C., T. Jenkins and G. Schatzmayr, 2019. Global mycotoxin occurrence in feed: A ten-year survey. Toxins, 11.
- Apeh, D.O., A.E. Bello, Y. Sunday, K.A. Apeh, M.A. Aiyegbuisi and Y. Ekeyi, 2024. Comprehensive review of mycotoxin exposure in Nigerian infants: Biomarker analysis and baby food contamination. Confluence Univ. J. Sci. Technol., 1: 92-100.
- Adetuniji, M.C., O.O. Atanda, C.N. Ezekiel, A.O. Dipeolu, S.V.A. Uzochukwu, J. Oyedepo and C.A. Chilaka, 2014. Distribution of mycotoxins and risk assessment of maize consumers in five agro-ecological zones of Nigeria. Eur. Food Res. Technol., 239: 287-296.
- Xu, R., E.G. Kiarie, A. Yiannikouris, L. Sun and N.A. Karrow, 2022. Nutritional impact of mycotoxins in food animal production and strategies for mitigation. J. Anim. Sci. Biotechnol., 13.
- Chen, X., M.F. Abdallah, X. Chen and A. Rajkovic, 2023. Current knowledge of individual and combined toxicities of aflatoxin B1 and fumonisin B1 in vitro. Toxins, 15.
- Cheng, K., J. Niu, X. Zheng, Y. Qiao and J. Zhang et al., 2023. Aflatoxin-B1-exposure-induced hepatic injury could be alleviated by polydatin through reducing oxidative stress, inhibiting inflammation and improving mitophagy. Toxics, 11.
- Turner, P.C., A.C. Collinosn, Y.B. Cheung, Y. Gong, A.J. Hall, A.M. Prentice and C.P. Wild, 2007. Aflatoxin exposure in utero causes growth faltering in Gambian infants. Int. J. Epidemiol., 36: 1119-1125.
- Nji, Q.N., O.O. Babalola, T.I. Ekwomadu, N. Nleya and M. Mwanza, 2022. Six main contributing factors to high levels of mycotoxin contamination in African foods. Toxins, 14.
- Visser, M.E., A. Schoonees, C.N. Ezekiel, N.P. Randall and C.E. Naude, 2020. Agricultural and nutritional education interventions for reducing aflatoxin exposure to improve infant and child growth in low- and middle-income countries. Cochrane Database Syst. Rev.
- Kimanya, M.E., B. de Meulenaer, D. Roberfroid, C. Lachat and P. Kolsteren, 2010. Fumonisin exposure through maize in complementary foods is inversely associated with linear growth of infants in Tanzania. Mol. Nutr. Food Res., 54: 1659-1667.
- Ingenbleek, L., M. Sulyok, A. Adegboye, S.E. Hossou and A.Z. Koné et al., 2019. Regional Sub-Saharan Africa total diet study in Benin, Cameroon, Mali and Nigeria reveals the presence of 164 mycotoxins and other secondary metabolites in foods. Toxins, 11.
- Wild, C.P. and Y.Y. Gong, 2010. Mycotoxins and human disease: A largely ignored global health issue. Carcinogenesis, 31: 71-82.
- Anih, D.C., A.K. Arowora, M.A. Abah and K.C. Ugwuoke, 2025. Biochemical effects of microplastics on human health: A comprehensive review. Sci. Int., 13: 27-34.
- Anih, D.C., O.E. Yakubu, A.K. Arowora, M.A. Abah and U.K. Chinekwu, 2025. Biochemical mechanisms of sleep regulation. Sci. Int., 13: 35-45.
- Anih, D.C., A.K, Arowora, M.A. Abah and K.C. Ugwuoke, 2025. Biochemically active metabolites of gut bacteria: Their influence on host metabolism, neurotransmission, and immunity. Sci. Int., 13: 46-57.
- Chinonso, A.D., A.A. Kayode, M.A. Adondua and U.K. Chinekwu, 2025. Biochemistry of traditional herbal compounds and their molecular targets. Pharmacogn. Rev., 19: 83-90.
- Arowora, A.K., I. Chinedu, D.C. Anih, A.A. Moses and K.C. Ugwuoke, 2022. Application of artificial intelligence in biochemistry and biomedical sciences: A review. Asian Res. J. Curr. Sci., 4: 302-312.
How to Cite this paper?
APA-7 Style
Anih,
D.C., Arowora,
K.A., Ugwuoke,
K.C., Abah,
M.A., Habibu,
B. (2025). Nutritional Modulation of Epigenetic Changes Induced by Mycotoxins: A Biochemical Perspective for At-Risk Populations in Africa. Science International, 13(1), 90-109. https://doi.org/10.17311/sciintl.2025.90.109
ACS Style
Anih,
D.C.; Arowora,
K.A.; Ugwuoke,
K.C.; Abah,
M.A.; Habibu,
B. Nutritional Modulation of Epigenetic Changes Induced by Mycotoxins: A Biochemical Perspective for At-Risk Populations in Africa. Sci. Int 2025, 13, 90-109. https://doi.org/10.17311/sciintl.2025.90.109
AMA Style
Anih
DC, Arowora
KA, Ugwuoke
KC, Abah
MA, Habibu
B. Nutritional Modulation of Epigenetic Changes Induced by Mycotoxins: A Biochemical Perspective for At-Risk Populations in Africa. Science International. 2025; 13(1): 90-109. https://doi.org/10.17311/sciintl.2025.90.109
Chicago/Turabian Style
Anih, David, Chinonso, Kayode Adebisi Arowora, Kenneth Chinekwu Ugwuoke, Moses Adondua Abah, and Bilyaminu Habibu.
2025. "Nutritional Modulation of Epigenetic Changes Induced by Mycotoxins: A Biochemical Perspective for At-Risk Populations in Africa" Science International 13, no. 1: 90-109. https://doi.org/10.17311/sciintl.2025.90.109

This work is licensed under a Creative Commons Attribution 4.0 International License.