Biochemical Mechanisms of Sleep Regulation
-
David Chinonso Anih


Department of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba State, Nigeria
Ojochenemi Ejeh YakubuDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba State, Nigeria
Adebisi Kayode AroworaDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba State, Nigeria
Moses Adondua AbahDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba State, Nigeria
Ugwuoke Kenneth ChinekwuDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba State, Nigeria
| Received 12 Apr, 2025 |
Accepted 18 Jun, 2025 |
Published 19 Jun, 2025 |
Sleep is a highly orchestrated neurophysiological process governed by intricate biochemical pathways involving neurotransmitters and hormones that modulate sleep architecture and circadian rhythmicity. This review presents a comprehensive synthesis of current molecular insights into the regulatory mechanisms of sleep, with a particular focus on the roles of γ-aminobutyric acid (GABA), serotonin, adenosine, acetylcholine, and norepinephrine in sleep stage modulation. It further explores the circadian and homeostatic functions of hormones, including melatonin, cortisol, orexins, leptin, and ghrelin, highlighting their integrative roles in linking neural circuitry with systemic physiological states. The article delineates how disruptions in these pathways underpin major sleep disorders, such as insomnia, narcolepsy, obstructive sleep apnea, restless leg syndrome, and circadian rhythm sleep-wake disorders, through mechanisms ranging from GABAergic deficits and HPA axis hyperactivation to orexin deficiency and clock gene dysregulation. By bridging neurochemical theory with clinical pathology, the review underscores the translational potential of targeting neuroendocrine circuits for therapeutic intervention. The findings advocate for the advancement of personalized sleep medicine through molecular diagnostics and emphasize the importance of circadian coherence in neurodegenerative and metabolic disease management. Collectively, this work consolidates multidisciplinary evidence into a cohesive framework, contributing to the evolving paradigm of sleep biology and its clinical relevance.
| Copyright © 2025 Anih et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Sleep is a fundamental, evolutionarily conserved biological process essential for sustaining neurocognitive performance, metabolic balance, and immune function. Globally, the burden of sleep-related disorders continues to rise, with profound consequences on mental health, cardiovascular integrity, and productivity. Over the past two decades, substantial progress has been made in elucidating the neurochemical basis of sleep, particularly the roles of neurotransmitters and hormones in orchestrating the sleep-wake cycle. Studies have established that γ-aminobutyric acid (GABA), serotonin, adenosine, acetylcholine, and norepinephrine mediate neural excitability and transitions between sleep stages1-5, while melatonin, cortisol, orexins, leptin, and ghrelin coordinate systemic signals with circadian cues6-9. Despite these insights, a comprehensive synthesis that bridges neurochemical mechanisms with the pathophysiology of sleep disorders remains limited. Previous reviews have often examined neurotransmitters or hormones in isolation, without integrating their dynamic interactions or exploring emerging therapeutic targets based on recent molecular findings. This review aims to address this gap by presenting an updated, integrative overview of the biochemical regulation of sleep, with a focus on neurotransmitters and hormones, and their relevance to sleep disorders. By consolidating current molecular evidence, this article seeks to inform future research directions and therapeutic approaches in sleep medicine.
NEUROTRANSMITTERS IN SLEEP REGULATION
Neurotransmitters are central to the neurobiological orchestration of sleep. Their synthesis, release, receptor binding, and degradation influence neuronal excitability, synaptic communication, and the homeostatic regulation of sleep cycles.
GABA
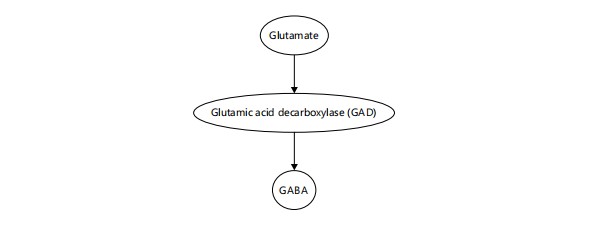
Principal inhibitory neurotransmitter: The GABA (γ-aminobutyric acid) is the chief inhibitory neurotransmitter in the CNS, playing a dominant role in sleep induction and maintenance by inhibiting arousal-promoting neurons in the hypothalamus and brainstem. The GABA is synthesized from glutamate through the catalytic action of the enzyme Glutamic Acid Decarboxylase (GAD), as illustrated in Fig. 1. This conversion is a critical biochemical pathway that governs the balance between excitatory and inhibitory signaling in the brain. By reducing glutamate levels and increasing GABA availability, this reaction facilitates the inhibition of wake-promoting neurons and supports the initiation and maintenance of sleep. The Ventrolateral Preoptic Nucleus (VLPO) releases GABA to inhibit wake-promoting centers such as the tuberomammillary nucleus, locus coeruleus, and raphe nuclei10. Melatonin enhances the effect of GABA by increasing the expression and sensitivity of GABAA receptors, particularly in the hypothalamus11. Sleep-promoting pharmacological agents, including benzodiazepines, zolpidem, and barbiturates, act as positive allosteric modulators of GABAA receptors, further validating the centrality of GABAergic transmission in sleep12.
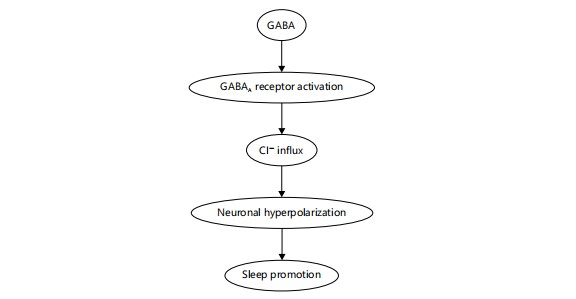
Upon release, GABA binds to GABAA receptors, leading to the influx of chloride ions (Cl–) into the postsynaptic neuron. This causes neuronal hyperpolarization, reducing neuronal excitability and facilitating sleep induction, especially during non-REM sleep. Figure 2 outlines this mechanistic pathway, visually demonstrating how GABAergic neurotransmission leads to CNS inhibition and ultimately supports sleep regulation12.
Serotonin
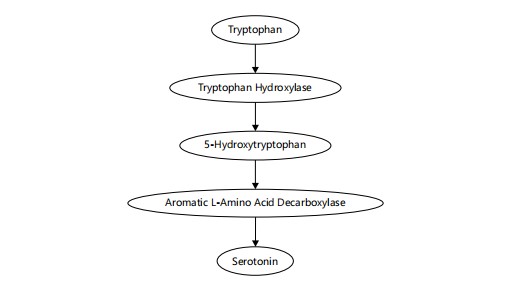
A multifaceted modulator: Serotonin (5-HT), derived from the amino acid tryptophan, is synthesized primarily in the raphe nuclei and influences both sleep initiation and circadian rhythm synchronization. It plays dual roles: While certain serotonergic pathways promote sleep onset, others may promote Biosynthesis of serotonin from tryptophan through the intermediate 5-hydroxytryptophan, catalyzed by tryptophan hydroxylase and Aromatic L-amino acid decarboxylase15 wakefulness depending on receptor subtype and regional activity. Additionally, serotonin is the precursor for melatonin synthesis in the pineal gland13. The SSRIs and other serotonergic agents significantly affect REM sleep patterns, sometimes reducing REM duration and altering sleep continuity14.
|
|
|
The essential amino acid tryptophan is first converted by tryptophan hydroxylase into 5-hydroxytryptophan (5-HTP). Then, Aromatic L-amino acid decarboxylase (AADC) catalyzes the decarboxylation of 5-HTP to produce serotonin (5-hydroxytryptamine, 5-HT). Figure 3 visually represents this biosynthetic pathway, emphasizing the enzymatic steps that are essential for serotonin production and its regulatory role in sleep-wake cycles15.
Adenosine
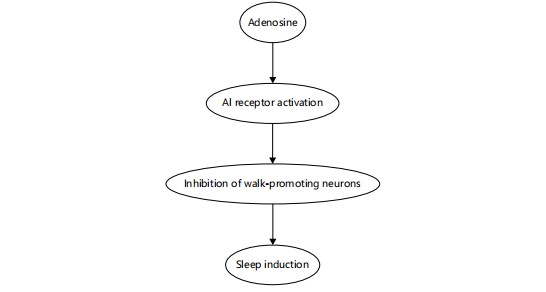
A homeostatic sleep factor: Adenosine accumulates during prolonged wakefulness as a byproduct of ATP metabolism and acts as a powerful homeostatic sleep inducer. It binds to A1 and A2A receptors in A process integral to sleep homeostasis. During periods of prolonged wakefulness, intracellular ATP is hydrolyzed to AMP, which is further converted to adenosine. Accumulating extracellular adenosine binds to A1 receptors in the brain, particularly in the basal forebrain, to inhibit wake-promoting neurons and promote sleep onset15 sleep-related regions such as the basal forebrain and inhibits wake-promoting neurotransmission. Studies have shown that the infusion of adenosine or A2A receptor agonists promotes NREM sleep, whereas A1 receptor knockout mice exhibit reduced sleep pressure15,16. Caffeine, a competitive antagonist of adenosine receptors, delays sleep onset and reduces total sleep time, highlighting adenosine’s critical sleep-promoting role17.

|
During prolonged neuronal activity or metabolic demand, ATP (Adenosine Triphosphate) is hydrolyzed to AMP (Adenosine monophosphate), which is subsequently converted into adenosine. As adenosine accumulates extracellularly, it binds to A1 receptors, especially in the basal forebrain, inhibiting wake-promoting neurons and promoting sleep onset. As illustrated in Fig. 4, adenosine is generated through the stepwise breakdown of ATP, first into AMP and subsequently into adenosine. This accumulation occurs progressively during sustained wakefulness and plays a crucial role in promoting sleep by inhibiting wake-active neurons via A1 receptors, particularly in the basal forebrain. The figure highlights the biochemical cascade from cellular energy depletion to extracellular adenosine build-up, emphasizing its homeostatic role in sleep regulation. This process has been well documented in both animal and human studies, supporting adenosine as a key sleep-inducing factor15,17.
Adenosine binds to A1 receptors in the brain, particularly in areas like the basal forebrain. Activation of these receptors leads to the inhibition of wake-promoting neurons, reducing arousal and leading to sleep induction. This mechanism plays a key role in regulating sleep pressure during extended periods of wakefulness. Beyond its accumulation, adenosine exerts its sleep-promoting effects through targeted neuromodulation. As depicted in Fig. 5, adenosine binds to A1 receptors located in key arousal-related brain regions, such as the basal forebrain. This receptor activation inhibits wake-promoting neurons, reducing cortical arousal and promoting the transition to sleep15,17. The figure complements the metabolic pathway outlined in Fig. 4 by illustrating the downstream neurophysiological effects of adenosine that underpin its homeostatic role in sleep regulation.
Acetylcholine
Regulator of REM sleep: Acetylcholine is synthesized in cholinergic neurons of the basal forebrain and pontine tegmentum. It is primarily associated with cortical arousal and REM sleep, where it facilitates desynchronized EEG activity and muscle atonia. The REM-on neurons in the Laterodorsally and Pedunculopontine Tegmental Nuclei (LDT/PPT) release acetylcholine during REM sleep episodes18. Anticholinergic drugs tend to suppress REM sleep, and cholinesterase inhibitors such as donepezil can increase REM density, demonstrating acetylcholine’s critical role in sleep architecture19.
|
This diagram illustrates how extracellular adenosine Activates A1 receptors, leading to inhibition of wake-promoting neurons in regions such as the basal forebrain and cortex. This neural suppression contributes to increased sleep pressure and facilitates sleep onset15,17
Norepinephrine
Modulator of arousal: Norepinephrine, secreted by the locus coeruleus, promotes wakefulness and vigilance. Its activity is highest during wakefulness, decreases during NREM sleep, and becomes nearly silent during REM sleep. However, recent studies suggest that phasic norepinephrine bursts during sleep might contribute to memory consolidation and synaptic homeostasis without fully awakening the brain, implying a nuanced role beyond simple arousal20,21. Dysregulation of noradrenergic tone is implicated in insomnia, PTSD, and other disorders involving disrupted sleep-wake transitions.
HORMONAL REGULATION OF SLEEP
Hormones act as systemic modulators of sleep by linking central neural circuits with peripheral physiological states. Their circadian patterns of secretion, influenced largely by light-dark cycles and metabolic cues, help regulate sleep timing, duration, and quality.
Melatonin
Circadian pacemaker: Melatonin, synthesized from serotonin in the pineal gland, is a central regulator of circadian rhythms. Its production is triggered by darkness and inhibited by light through input from the Suprachiasmatic Nucleus (SCN), which transmits light information from the retina via the retinohy pothalamic tract22. The enzyme Hydroxyindole-O-methyltransferase (HIOMT) catalyzes the final step in melatonin synthesis. Once secreted, melatonin binds to MT1 and MT2 receptors in the SCN to induce sleepiness and synchronize the circadian clock23. The biochemical pathway underlying this conversion of serotonin into melatonin is illustrated in Fig. 6, which highlights the roles of N-acetyltransferase (NAT) and Hydroxyindole-O-methyltransferase (HIOMT) in this essential circadian regulatory mechanism22-24. Studies have shown that exogenous melatonin supplementation can improve sleep onset latency in individuals with delayed sleep-wake phase disorder and in elderly populations with reduced endogenous production24.
In the pineal gland, serotonin is first acetylated by N-acetyltransferase (NAT) to form N-acetyl serotonin. This intermediate is then methylated by Hydroxyindole-O-methyltransferase (HIOMT), producing melatonin. The secretion of melatonin is strongly regulated by the light-dark cycle and plays a critical role in promoting sleep22-24.

|
Cortiso
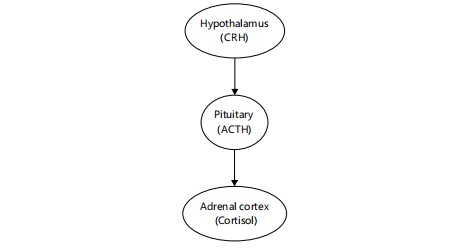
Arousal hormone: Cortisol is secreted by the adrenal cortex in a circadian pattern regulated by the Hypothalamic-pituitary-adrenal (HPA) axis. It peaks in the early morning hours, promoting alertness and energy mobilization, and gradually declines throughout the day. Stress-induced HPA axis activation elevates nocturnal cortisol levels, which are associated with sleep fragmentation and reduced slow-wave sleep25,26. Figure 7 illustrates this hormonal cascade, beginning with the hypothalamus releasing Corticotropin-releasing Hormone (CRH), which stimulates the anterior pituitary to release Adrenocorticotropic Hormone (ACTH), ultimately leading to cortisol secretion by the adrenal cortex7,26,27. This dysregulation is frequently implicated in insomnia and mood disorders, where hyperactivity of the HPA axis perpetuates a cycle of poor sleep and heightened arousal7,25,27.
Orexins (Hypocretins)
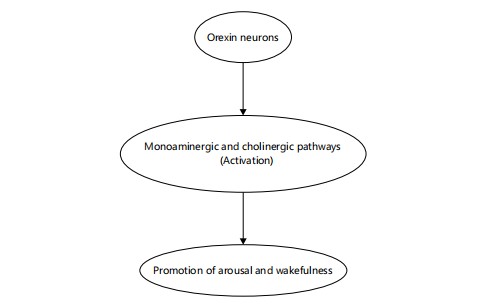
Integrators of wakefulness: Orexin-A and Orexin-B, neuropeptides synthesized in the lateral hypothalamus, are essential for sustaining wakefulness and preventing inappropriate transitions into REM sleep. These peptides activate monoaminergic and cholinergic arousal systems and stabilize sleep-wake transitions28. Figure 8 illustrates this neurochemical cascade, where orexin neurons enhance the activity of downstream arousal circuits, ultimately promoting and maintaining wakefulness28-30. Orexin deficiency is strongly implicated in narcolepsy, highlighting its vital role in sleep-wake regulation28,29. Deficiency in orexin neurons, due to autoimmune or genetic factors, is the primary cause of narcolepsy type 129. Orexin receptor antagonists, such as suvorexant, have been developed as novel therapeutics for insomnia, demonstrating the translational potential of targeting this system30.
Leptin and Ghrelin
Metabolic hormones influencing sleep: Leptin, produced by adipose tissue, promotes satiety and has been shown to enhance sleep, possibly by inhibiting orexin neurons31. Ghrelin, secreted by the stomach, stimulates hunger and may antagonize leptin’s effects on sleep. Sleep deprivation leads to decreased leptin and increased ghrelin levels, contributing to increased appetite and weight gain32. These hormones are dysregulated in obesity-related sleep disorders such as obstructive sleep apnea (OSA), further linking metabolic and sleep homeostasis33.
|
|
BIOCHEMICAL DYSREGULATION IN SLEEP DISORDERS
Sleep disorders often arise from disruptions in neurotransmitter and hormonal systems. Understanding these molecular pathologies is essential for the development of targeted therapies.
Insomnia
GABA and cortisol imbalance: Insomnia, the most prevalent sleep disorder, is often associated with hyperarousal due to reduced GABAergic inhibition and excessive HPA axis activation. Neuroimaging studies have shown decreased GABA levels in the anterior cingulate cortex of insomniacs34. Concurrently, elevated nocturnal cortisol impairs sleep initiation and maintenance. Pharmacological interventions such as benzodiazepines, zolpidem, and eszopiclone target GABAA receptors to restore inhibitory tone and promote sleep35. Cognitive-behavioral Therapy for insomnia (CBT-I) has also been shown to reduce cortisol levels and improve sleep quality36.
Narcolepsy
Orexin deficiency: Narcolepsy Type 1 is characterized by excessive daytime sleepiness, cataplexy, and disturbed nocturnal sleep. The underlying cause is a loss of orexin-producing neurons, often due to autoimmune mechanisms triggered by HLA-DQB1*06:02 positivity and possibly viral infections37,38. Current treatments include modafinil, sodium oxybate, and orexin receptor agonists in development, such as danavorexton, which aim to restore orexin signaling39.
Sleep apnea and circadian hormonal disruption: The OSA is associated with intermittent hypoxia, fragmented sleep, and altered secretion of circadian hormones. Melatonin levels are often reduced in OSA patients, while the normal cortisol rhythm becomes blunted, potentially exacerbating cardiovascular and metabolic complications40. The CPAP therapy has been shown to restore hormonal rhythms, reduce oxidative stress, and improve cognitive performance in patients with moderate to severe OSA41.
Restless leg syndrome (RLS) and dopaminergic dysregulation: The RLS is a sensorimotor disorder linked to dysfunction in the dopaminergic system and iron deficiency. Iron is a necessary cofactor for tyrosine hydroxylase, the enzyme that catalyzes dopamine synthesis. The MRI studies have shown reduced iron concentrations in the substantia nigra of RLS patients42.
Although primarily dopaminergic, RLS may also reflect broader circadian misalignments. Dopamine agonists like pramipexole and ropinirole are effective in reducing RLS symptoms, although long-term use can lead to augmentation, necessitating careful clinical management43.
Circadian rhythm sleep-wake disorders and clock gene disruption: Circadian Rhythm Sleep-Wake Disorders (CRSWDs) result from desynchronization between endogenous biological rhythms and environmental time cues. At the molecular level, these disorders are often linked to disruptions in the expression or function of core clock genes such as CLOCK, BMAL1, PER, and CRY, which regulate rhythmic transcriptional cycles across the central nervous system and peripheral tissues. Dysregulation of these genes impairs the sleep-wake cycle, leading to conditions like delayed sleep phase disorder, shift work disorder, and non-24 hrs sleep-wake rhythm disorder44. Emphasized that clock gene integrity is critical to maintaining circadian coherence, as these genetic networks influence melatonin secretion, core body temperature rhythms, and hormonal oscillations44. The CRSWDs have been associated with increased risk of mood disturbances, metabolic disorders, and reduced cognitive performance. Therapeutic approaches such as light therapy, melatonin administration, and sleep scheduling have been shown to improve entrainment by targeting the circadian system44.
Neurodegenerative sleep disorders and circadian clock impairment: In neurodegenerative diseases such as Alzheimer’s and Parkinson’s, sleep disturbances are strongly linked to circadian rhythm dysfunction. Degeneration of the Suprachiasmatic nucleus (SCN), the central pacemaker, along with reduced expression of circadian genes, leads to fragmented sleep patterns, blunted melatonin rhythms, and dysregulated hormonal outputs. Yakubu et al.44 highlighted the role of circadian disruption in exacerbating both sleep and cognitive symptoms in these diseases, suggesting that neurodegeneration itself may impair molecular timekeeping44. In Alzheimer’s, diminished melatonin secretion and SCN atrophy correlate with increased nocturnal activity and daytime sleepiness. In Parkinson’s, circadian phase shifts and REM sleep behavior disorder reflect dopaminergic degeneration affecting circadian synchrony. Circadian-based interventions, such as timed light exposure and melatonin therapy, can improve sleep architecture and may potentially slow cognitive decline when administered early44.
CONCLUSION
Sleep regulation is governed by a complex interplay of neurochemical systems, with key roles played by GABA, serotonin, adenosine, melatonin, and orexins. Disruptions in these pathways contribute to the pathophysiology of various sleep disorders. Integrating molecular insights with established neurobiological frameworks supports the advancement of targeted, precision-based therapies. Continued interdisciplinary research is essential to unravel the intricate mechanisms of sleep and to enhance clinical outcomes in associated neuropsychiatric and metabolic conditions.
SIGNIFICANCE STATEMENT
Sleep disorders affect millions worldwide, yet the underlying biochemical mechanisms governing sleep regulation remain incompletely understood, hindering the development of targeted therapies. This review elucidates the complex interplay between neurotransmitters (e.g., GABA, serotonin, adenosine) and hormones (e.g., melatonin, cortisol, orexins) in modulating sleep stages, circadian rhythms, and the pathophysiology of common sleep disorders. By integrating molecular, neurochemical, and hormonal evidence, the article provides a cohesive framework that advances understanding of sleep biology. These insights not only inform therapeutic strategies for insomnia, narcolepsy, and sleep apnea but also support the growing field of personalized sleep medicine. Furthermore, the findings have broader implications for managing neuropsychiatric and metabolic conditions, emphasizing the interdisciplinary relevance of sleep research in public health and clinical practice.
REFERENCES
- St-Onge, M.P., A. Mikic and C.E. Pietrolungo, 2016. Effects of diet on sleep quality. Adv. Nutr., 7: 938-949.
- Yu, Q., Q. Guo, S. Jin, C. Gao, P. Zheng, D.P. Li and Y. Wu, 2023. Melatonin suppresses sympathetic vasomotor tone through enhancing GABAA receptor activity in the hypothalamus. Front. Physiol., 14.
- Cools, R., K. Nakamura and N.D. Daw, 2011. Serotonin and dopamine: Unifying affective, activational, and decision functions. Neuropsychopharmacology, 36: 98-113.
- Huang, Z.L., Y. Urade and O. Hayaishi, 2011. The role of adenosine in the regulation of sleep. Curr. Top. Med. Chem., 11: 1047-1057.
- Saper, C.B., P.M. Fuller, N.P. Pedersen, J. Lu and T.E. Scammell, 2010. Sleep state switching. Neuron, 68: 1023-1042.
- Lian, X., Z. Liu, Z. Gan, Q. Yan and L. Tong et al., 2025. Targeting the glymphatic system to promote α-synuclein clearance: A novel therapeutic strategy for Parkinson’s disease. Neural Regener. Res., 21: 233-247.
- Li, Y., L. Lu and I.P. Androulakis, 2024. The physiological and pharmacological significance of the circadian timing of the HPA axis: A mathematical modeling approach. J. Pharm. Sci., 113: 33-46.
- Bjorness, T.E. and R.W. Greene, 2024. Orexin-mediated motivated arousal and reward seeking. Peptides, 180.
- Knutson, K.L., K. Spiegel, P. Penev and E. van Cauter, 2007. The metabolic consequences of sleep deprivation. Sleep Med. Rev., 11: 163-178.
- Valero, M., T.J. Viney, R. Machold, S. Mederos and I. Zutshi et al. 2021. Sleep down state-active ID2/Nkx2.1 interneurons in the neocortex. Nat. Neurosci., 24: 401-411.
- Holbrook, A.M., R. Crowther, A. Lotter, C. Cheng and D. King, 2000. Meta-analysis of benzodiazepine use in the treatment of insomnia. Can. Med. Assoc. J., 162: 225-233.
- Scammell, T.E., E. Arrigoni and J.O. Lipton, 2017. Neural circuitry of wakefulness and sleep. Neuron, 93: 747-765.
- Monti, J.M., 2011. Serotonin control of sleep-wake behavior. Sleep Med. Rev., 15: 269-281.
- Wichniak, A., A. Wierzbicka and W. Jernajczyk, 2012. Sleep and antidepressant treatment. Curr. Pharm. Des., 18: 5802-5817.
- Porkka-Heiskanen, T., R.E. Strecker, M. Thakkar, A.A. Bjørkum, R.W. Greene and R.W. McCarley, 1997. Adenosine: A mediator of the sleep-inducing effects of prolonged wakefulness. Science, 276: 1265-1268.
- Basheer, R., R.E. Strecker, M.M. Thakkar and R.W. McCarley, 2004. Adenosine and sleep-wake regulation. Prog. Neurobiol., 73: 379-396.
- Fredholm, B.B., K. Bättig, J. Holmén, A. Nehlig and E.E. Zvartau, 1999. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol. Rev., 51: 83-133.
- McCarley, R.W., 2007. Neurobiology of REM and NREM sleep. Sleep Med., 8: 302-330.
- Mihaylova, R., V.T. Angelova, J. Tchekalarova, D. Atanasova, P. Ivanova and R. Simeonova, 2024. Tailored melatonin- and donepezil-based hybrids targeting pathognomonic changes in Alzheimer’s disease: An in vitro and in vivo investigation. Int. J. Mol. Sci., 25.
- Andersen, L.P.H., I. Gögenur, J. Rosenberg and R.J. Reiter, 2016. The safety of melatonin in humans. Clin. Drug Invest., 36: 169-175.
- Sara, S.J., 2009. The locus coeruleus and noradrenergic modulation of cognition. Nat. Rev. Neurosci., 10: 211-223.
- Czeisler, C.A., J.S. Allan, S.H. Strogatz, J.M. Ronda and R. Sánchez et al., 1986. Bright light resets the human circadian pacemaker independent of the timing of the sleep-wake cycle. Science, 233: 667-671.
- Winrow, C.J., A.L. Gotter, C.D. Cox, S.M. Doran and P.L. Tannenbaum et al., 2011. Promotion of sleep by Suvorexant-A novel dual orexin receptor antagonist. J. Neurogenet., 25: 52-61.
- Ferracioli-Oda, E., A. Qawasmi and M.H. Bloch, 2013. Meta-analysis: Melatonin for the treatment of primary sleep disorders. PLoS One, 8.
- Born, J. and H.L. Fehm, 2000. The neuroendocrine recovery function of sleep. Noise Health, 2: 25-37.
- Meerlo, P., A. Sgoifo and D. Suchecki, 2008. Restricted and disrupted sleep: Effects on autonomic function, neuroendocrine stress systems and stress responsivity. Sleep Med. Rev., 12: 197-210.
- Vgontzas, A.N. and G.P. Chrousos, 2002. Sleep, the hypothalamic-pituitary-adrenal axis, and cytokines: Multiple interactions and disturbances in sleep disorders. Endocrinol. Metab. Clin. N. Am., 31: 15-36.
- Besing, G.L.K., E.K.S. John, C.V. Potesta, M.J. Gallagher and C. Zhou, 2022. Artificial sleep-like up/down-states induce synaptic plasticity in cortical neurons from mouse brain slices. Front. Cell. Neurosci., 16.
- Mignot, E., 1998. Genetic and familial aspects of narcolepsy. Neurology, 50: S16-S22.
- Morin, C.M. and D.J. Buysse, 2024. Management of Insomnia. N. Engl. J. Med., 391: 247-258.
- Pan, W. and A.J. Kastin, 2014. Leptin: A biomarker for sleep disorders? Sleep Med. Rev., 18: 283-290.
- Spiegel, K., E. Tasali, P. Penev and E. van Cauter, 2004. Brief communication: Sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite. Ann. Intern. Med., 141: 846-850.
- Punjabi, N.M., 2008. The epidemiology of adult obstructive sleep apnea. Proc. Am. Thoracic Soc., 5: 136-143.
- Winkelman, J.W., O.M. Buxton, J.E. Jensen, K.L. Benson, S.P. O'Connor, W. Wang and P.F. Renshaw, 2008. Reduced brain GABA in primary insomnia: Preliminary data from 4T proton magnetic resonance spectroscopy (1H-MRS). Sleep, 31: 1499-1506.
- Jansson-Fröjmark, M., C. Sandlund and A. Norell-Clarke, 2023. Paradoxic intention as an adjunct treatment to cognitive behavioral therapy for insomnia. Sleep Med. Clin., 18: 9-19.
- Tang, N.K.Y. and A.G. Harvey, 2006. Altering misperception of sleep in insomnia: Behavioral experiment versus verbal feedback. J. Consulting Clin. Psychol., 74: 767-776.
- Cvetkovic-Lopes, V., L. Bayer, S. Dorsaz, S. Maret and S. Pradervand et al., 2010. Elevated tribbles homolog 2-specific antibody levels in narcolepsy patients. J. Clin. Invest., 120: 713-719.
- Mahlios, J., A.K. de la Herrán-Arita and E. Mignot, 2013. The autoimmune basis of narcolepsy. Curr. Opin. Neurobiol., 23: 767-773.
- Allen, R.P., D.L. Picchietti, D. Garcia-Borreguero, W.G. Ondo and A.S. Walters et al., 2014. Restless legs syndrome/Willis-Ekbom disease diagnostic criteria: Updated International Restless Legs Syndrome Study Group (IRLSSG) consensus criteria-history, rationale, description, and significance. Sleep Med., 15: 860-873.
- Macey, P.M., R. Kumar, M.A. Woo, E.M. Valladares, F.L. Yan-Go and R.M. Harper, 2008. Brain structural changes in obstructive sleep apnea. Sleep, 31: 967-977.
- Babu, A.R., J. Herdegen, L. Fogelfeld, S. Shott and T. Mazzone, 2005. Type 2 diabetes, glycemic control, and continuous positive airway pressure in obstructive sleep apnea. Arch. Intern. Med., 165: 447-452.
- Winkelman, J.W., J.A. Berkowski, L.M. DelRosso, B.B. Koo and M.T. Scharf et al., 2025. Treatment of restless legs syndrome and periodic limb movement disorder: An American Academy of Sleep Medicine systematic review, meta-analysis, and GRADE assessment. J. Clin. Sleep Med., 21: 153-199.
- Winkelman, J.W., J.A. Berkowski, L.M. DelRosso, B.B. Koo and M.T. Scharf et al., 2025. Treatment of restless legs syndrome and periodic limb movement disorder: An American Academy of Sleep Medicine clinical practice guideline. J. Clin. Sleep Med., 21: 137-152.
- Yakubu, O.E., J.I. Umaru, S.M.C. Udeh, E.M. Ale and T.E. Iorkuse, 2022. The circadian clock: An essential biological rhythm. Asian J. Res. Biosci., 4: 96-101.
How to Cite this paper?
APA-7 Style
Anih,
D.C., Yakubu,
O.E., Arowora,
A.K., Abah,
M.A., Chinekwu,
U.K. (2025). Biochemical Mechanisms of Sleep Regulation. Science International, 13(1), 35-45. https://doi.org/10.17311/sciintl.2025.35.45
ACS Style
Anih,
D.C.; Yakubu,
O.E.; Arowora,
A.K.; Abah,
M.A.; Chinekwu,
U.K. Biochemical Mechanisms of Sleep Regulation. Sci. Int 2025, 13, 35-45. https://doi.org/10.17311/sciintl.2025.35.45
AMA Style
Anih
DC, Yakubu
OE, Arowora
AK, Abah
MA, Chinekwu
UK. Biochemical Mechanisms of Sleep Regulation. Science International. 2025; 13(1): 35-45. https://doi.org/10.17311/sciintl.2025.35.45
Chicago/Turabian Style
Anih, David, Chinonso, Ojochenemi Ejeh Yakubu, Adebisi Kayode Arowora, Moses Adondua Abah, and Ugwuoke Kenneth Chinekwu.
2025. "Biochemical Mechanisms of Sleep Regulation" Science International 13, no. 1: 35-45. https://doi.org/10.17311/sciintl.2025.35.45

This work is licensed under a Creative Commons Attribution 4.0 International License.